Mitochondrial Inner Membrane Laboratory (MIM-Lab)
All living organisms depend on the ability to form membrane systems using phospholipids as membrane building blocks. The bacterial plasma membrane and mitochondrial inner membranes contain the unique diphosphatidyl glycerolipid cardiolipin. While the overall process of cardiolipin synthesis is known in terms of substrates, products and enzymes, detailed molecular insights into this pathway are limited, primarily due to the complete lack of structural knowledge of the enzymes. This gap also precludes fundamental understanding of mutations in cardiolipin-forming enzymes that cause severe diseases in affected patients. The goal of the MIM-Lab is to dissect the molecular nature of bacterial and mitochondrial cardiolipin synthases in order to understand how structural features dictates their functional activities.
We apply basic molecular biology methods such as cloning, protein expression in bacteria and mammalian systems and protein purification and we use cryo-electron microscopy for structure determination. Likewise, we also extract proteins and protein complexes from native tissue using differential centrifugation and density sedimentation methods.
The MIM-Lab is funded by the Novo Nordisk Foundation through a Hallas-Møller Emerging Investigator grant.
If our research and projects are any of your interest or you have any questions please get in touch via email (rkf@mbg.au.dk) or come by our lab on 5th floor in building 1874.
Mitochondria of eukaryotic cells are unique cell organelles enclosed by a double membrane system – the outer and the inner mitochondrial membrane. The inner mitochondrial membrane is a hotspot for bioenergetic protein complexes such as respiratory complexes of the electron transport chain and ATP synthase, which regenerates ATP for all cellular processes. Cardiolipin phospholipids are normally only found in the inner mitochondrial membrane where they serve to bind and stabilize protein complexes as well as to induce membrane curvature. In most mitochondria, cardiolipin is synthesized from phosphatidylglycerol and CDP-diacylglycerol, catalyzed by a small integral membrane protein called cardiolipin synthase. Currently, no experimental structures exist of this protein thus detailed insights into its function are lacking. However, by the use of AlphaFold, human cardiolipin synthase is believed to have six transmembrane helices characteristic of the CDP-AP protein family.
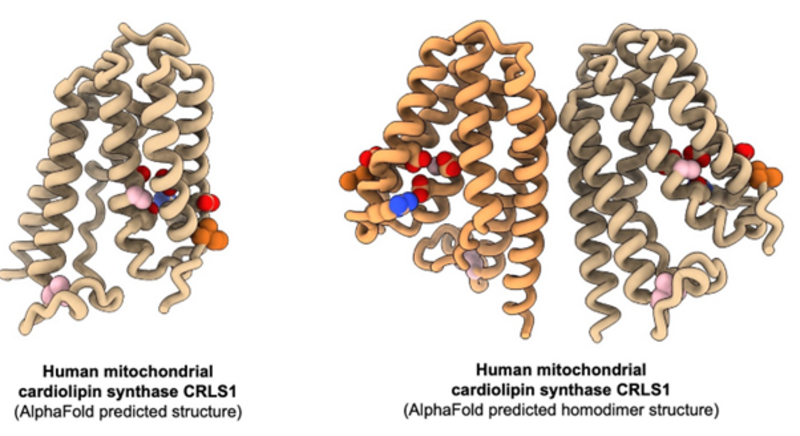
In this project we want to express and purify human cardiolipin synthase for structure determination by cryo-EM and activity assays. The structure will answer questions on the oligomeric state (monomer or dimer) and by adding individual substrates we will learn how the protein binds and recognizes the correct substrates for cardiolipin synthesis. From activity assays we can test which substrates the protein is active on, in particular how the acyl chain compositions influence the activity towards cardiolipin synthesis.
A longer-term goal for this project is to isolate intact mitochondria from native tissue and extract cardiolipin synthase from the inner mitochondrial membrane. By this approach we aim to map potential protein interaction partners and potentially see if the cardiolipin synthase engages in high molecular weight complexes. These complexes will be characterized and analyzed by a combination of cryo-EM, mass spectrometry and MD simulations.
Given the kinship of Gram-negative bacteria and mitochondria, they share many features including the presence of cardiolipin phospholipids in the membrane. In the bacterium Escherichia coli, cardiolipin locates to regions of high membrane curvature while also interacting with membrane proteins. However, the pathway for cardiolipin synthesis is rather different from the mitochondrial situation. Thus, in E. coli it is two molecules of phosphatidylglycerol that are condensed to form cardiolipin with release of one glycerol. The cardiolipin synthase responsible for this reaction belongs to the phospholipase D family of proteins, which are completely different than the mitochondrial cardiolipin synthase. Again, no experimental structures are available of the bacterial cardiolipin synthases, but using AlphaFold to predict their structures, we see no immediate transmembrane structural elements suggesting the protein to be a peripheral membrane protein. Furthermore, E. coli has a total of three genes encoding cardiolipin synthases, which seem to work independently under different conditions in the cell.
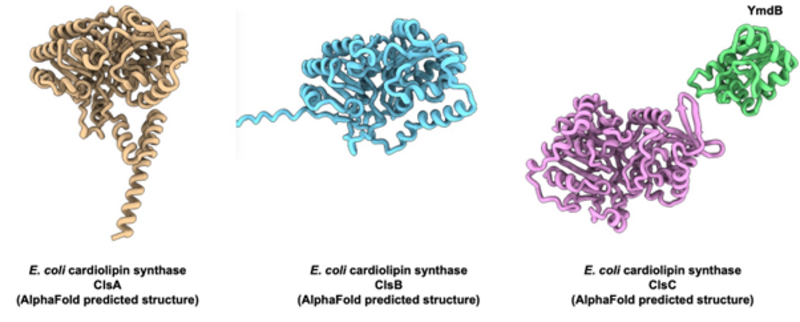
In this project we first want to express and purify the three different cardiolipin synthases from E. coli to establish activity assays where we can measure the catalytic differences between the three proteins, such as substrate specificity, kinetic parameters, etc. Second, we will determine the structures of the proteins reconstituted on lipid nanodiscs to mimic their peripheral membrane protein nature. By this we aim to understand how these proteins are able to extract substrates from the membrane and insert the product cardiolipin again.
Human parasites like Trypanosoma brucei and Toxoplasma gondii are single-celled eukaryotes which cause diseases in humans like African sleeping sickness and toxoplasmosis. Their mitochondria have very different morphologies compared to human mitochondria and the bioenergetic protein complexes that reside in the parasite mitochondria are highly diverged and adapted to exist in the unique membrane compartments. Studies have reported that mitochondrial cardiolipin synthases in the parasites are nothing like the canonical equivalents in human mitochondria, but instead resemble the bacterial cardiolipin synthases by their phospholipase D fold. This opens many interesting questions on mitochondrial evolution as well as basic biochemical questions on how these enzymes work in comparison to bacterial and other mitochondrial enzymes.
In this project, we first want to express and purify parasite cardiolipin synthases to assess their catalytic activities in comparison to bacterial and canonical mitochondrial enzymes. Here we will test lipid dependencies of the parasite enzymes as well as substrate specificities. Second, we will determine structures by cryo-EM and make comparative analyses to structures of bacterial structures to find differences and similarities for potential exploitation in drug development that can be tested in live parasites.

