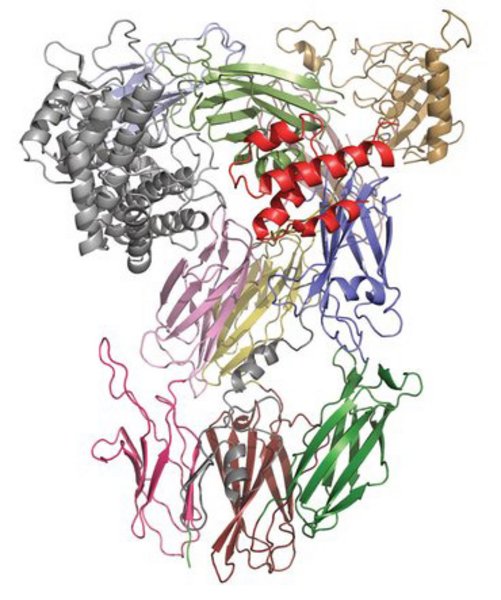
The research group determines crystal structures of large single proteins, protein-protein complexes, and protein-RNA complexes, and we have determined several structures in the 60-350 kDa range. We validate our structures with biophysical and functional experiments. In addition we also use small angle x-ray scattering to study the molecules in solution.
Our projects are focused on studying innate immunity, in particular how it recognizes danger signal on pathogens or damaged self-tissue and the response to the perception of such danger signals. An important component in the innate immune system is the complement system, formed by more than 50 soluble and membrane bound proteins. The soluble proteins are present in extracellular fluids, while the membrane proteins are located in membranes surrounding many of our cells, where they transmit the extracellular danger signal into intracellular signaling, protect our own cells from complement, and contributes to clearance of pathogens and host cells targeted by complement opsonization. We have determined structures of the key proteins C3, C4, and C5 in a variety of functional states, and hereby been able to propose how these proteins are proteolytically cleaved by enzymes assembled and activated in response to danger signals (see figure).
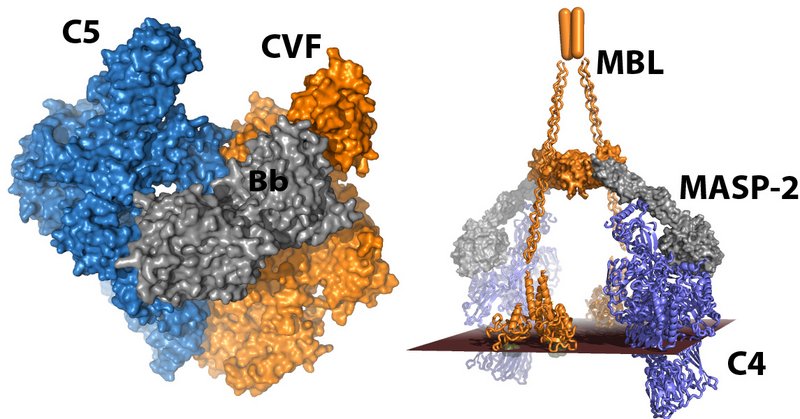
Figure. Models of the very large proteolytic complexes cleaving C3, C4 and C5. Left, A model of C5 (blue) bound to the cobra venom factor (orange) and the catalytically active serine protase Bb (grey). The complement convertases probably recognises their substrates C3 and C5 in a manner similar to this, see Laursen et al (2011) EMBO J. 30, 606-16. Right, Our model for the giant lectin pathway MBL·MASP-2 complex (orange and grey) cleaving its substrate C4 (blue), see Kidmose et al. (2012), PNAS 109, 15425-30. The red surface represent the glycan recognised by MBL, whereby MASP-2, the active serine protease, becomes active. Click figure for enlargement.
The innate immune system acts as a first line of defense against pathogens and in addition stimulates the adaptive immune system. It also plays an essential role in clearing apoptotic and necrotic cells thereby minimizing auto-immunity. Unfortunately, in a number of major disease conditions like diabetes, arthritis, cancer, Alzheimers disease, and sepsis the innate immune system contributes to an excessive or chronical inflammatory response. It is therefore of considerable interest to be able to selectively control these undesirable effects of innate immunity. The long term aim of our research is therefore also to develop selective inhibitors of the complement and other branches of innate immunity.
Our work within innate immunity is generously funded by a Hallas-Møller stipend to Gregers Rom Andersen from the Novo-Nordisk foundation www.novonordiskfonden.dk, the LUNA nanomedicine center www.nanomedicine.au.dk, the Lundbeck foundation www.lundbeckfonden.dk.
Metoder
Et projekt starter ofte med, at den studerende kloner genet for et protein og indsætter det i en ekspressionsvektor. Da vi arbejder med både extracellulære og membranbunde proteiner, udtrykker vi dem i bakterier, gær, insekt og mammale celler. De sidste er de bedste til at danne disulfidbroer og glykosylere proteinerne. Herefter oprenses de til høj renhed, før de krystalliseres. Nogle af proteinere fra komplementsystemet findes i høje koncentrationer i plasma, så disse kan oprenses direkte fra plasma.
Vi laver også funktionelle forsøg med f.eks. ITC og biokemiske assays, bl.a. for at checke at proteinerne er i en aktiv konformation. Vi får de rene proteiner eller komplekser mellem proteiner til at krystallisere ved at blande proteinopløsningen med diverse salte og organiske polymerer, som binder vand. Når krystaller er dannet, beskydes de med røntgenstråling, og ud fra det dannede diffraktionsmønster kan vi bestemme strukturen af proteinerne i krystallen. En vigtig efterfølgende fase er at bruge strukturen til at forklare kendte funktionelle data. Kan vi ikke krystallisere proteinet, beskyder vi proteinet i opløsning (SAXS), hvorved vi kan beregne formen af proteinet, men ikke detaljerede atomare modeller.
Et kortere forskningsprojekt i gruppen (10-20 ECTS) vil normalt kun omfatte en mindre del af hele arbejdsgangen fra gen til struktur, f.eks. at oprense et-to proteiner og forsøge at karakterisere og krystallisere disse. I specialer (60 ECTS) vil indgå større dele af arbejdsgangen, og dette kan lede til strukturbestemmelse af mindre proteiner. I ph.d.-projekter vil den studerende møde alle aspekter fra konstruktion af expressionssystemer til den endelige strukturanalyse. Vores ph.d.-studerende bruger normalt 10-20% af tiden på computerbaseret krystallografi og strukturanalyse, mens resten af tiden bruges på laboratorieforsøg med henblik på at frembringe, isolere, karakterisere og krystallisere proteiner og deres komplekser.


Read the article: Complement Structure (from Labtimes.org) (PDF-file).