Since 1993 we have been engaged with scientific studies on the proteins associated with the milk fat globule membrane (MFGM). Milk fat/lipid globules (size: 0.1 to 15 ?m) carries the lipid in milk and is a result of a secretion process directing newly intracellular synthesized lipid from the smooth ER of the mammary epithelial cell to this sophistically biological nourishing fluid. The MFGM serves to bring these hydrophobic molecules into “solution”.
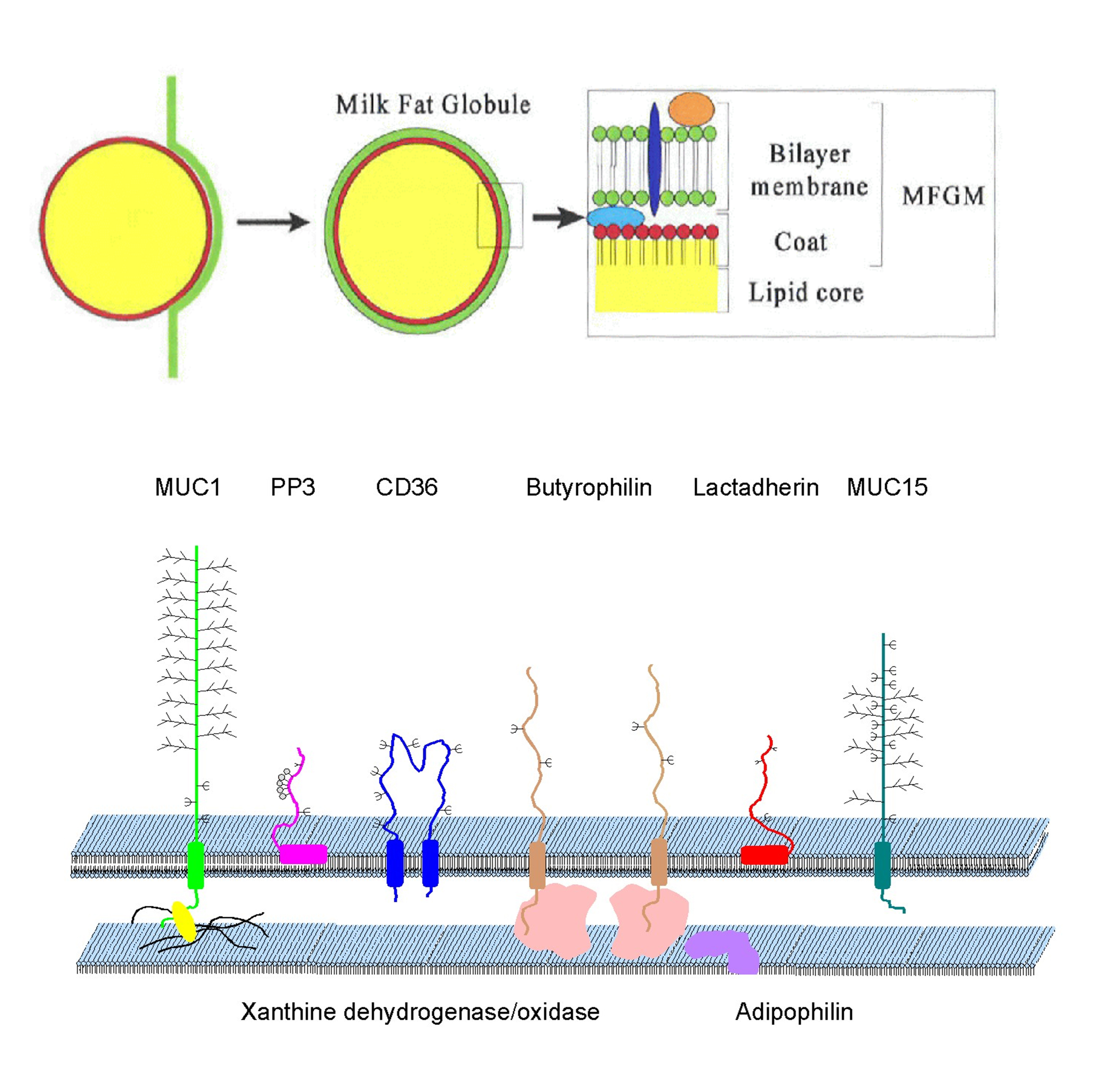
The bovine system constitutes about nine major MFGM proteins. Eight of them are depicted on the drawn model of MFGM on figure 1.
A vast amount of literature has been published concerning the appearance, structure, biological and nutritional role of individual MFGM components as well as their physiological impact in concert. Below is given a short description of a selection of MFGM proteins that we have studied. Few references are given, alternatively they might be found in Rasmussen, 2009, or in reviews like Mather, 2000; Dewettinck et al., 2008; Cavaletto et al., 2008. Many researchers have put a lot of effort into describing the significance and functionality of MFGM proteins. It is however meaningless to make this short piece fully referenced. I apologize to those not cited.
Figure 1. Schematic drawing of section process and model of the membrane of a newly secreted milk fat globule (sizes are not proportional). Milk triacylglycerides are located inside (below) surrounded by a unilaminar phospholipid membrane. The bilayered phospholipid membrane, with transmembrane proteins (MUC1, MUC15, Butyrophilin, and CD36) and peripheral bound proteins (Lacadherin and PP3), is a result of the envelopment process (top) taking place at the apical membrane of the milk producing cell. The intermembrane space is occupied with the cytosolic parts of the transmembrane proteins and “coat”-proteins (Xanthine oxidoreductase and Adipophilin). (Rasmussen et al., 2002). Click to enlarge photo.
The selective physical extracellular membrane that covers many epithelial surfaces has mucins as major constituents. Milk mucins are mainly MUC1, MUC4 (most likely equivalent to MUCX), and MUC15 (Pallesen et al., 2001; Zhang et al., 2005; Pallesen et al., 2002).
Most knowledge has been collected for MUC1, and in milk the attached negatively charged sialic acids most probably helps to prevent clogging and clumping of MFGs/milk and to keep ducts and alveoli open (Patton et al., 1995; Patton, 1999). The information about MUC4 and MUC15 are limited, the physiological role of MUC15 has, however, been addressed (Pallesen et al., 2007; Pallesen et al., 2008). Interestingly, MUC1 of consumed milk may also serve as protection against pathogenic bacteria, operating as bait and thereby decoying them away from the epithelium of the gastrointestinal tract (see e.g. Parker et al., 2010).

Figure 2. Schematic representation of structural characteristics known for milk mucins (MUC1, MUC4, and MUC15). SP, signal sequences; PTS, proline threonine and serine rich; EGF, epidermal growth factor; TM, transmembrane; CYT, cytosolic, VNTR, variable number of tandem repeats, VWFD, von Willebrand factor domain. See original literature for description of SEA, NIDO, and AMOP domains. Figure: Jan Trige Rasmussen. Click to enlarge figure.
Bovine milk xanthine oxidoreductase (XOD) was already isolated and studied in a relative pure form in 1924. In the following period it became one of the most thoroughly studied enzymes. After 62 years the complete amino acid sequences (1331 aa, 147 kDa) was published (Berglund et al., 1996). XOD contributes to purine degradation by oxidizing hypoxanthine to xanthine and subsequently to uric acid, but might also be linked to other activities and physiological roles (e.g. Puddu et al., 2008).
It has been proposed that milk XOD may play a protective antibacterial role in the digestive system of neonates by its capacity to generate reactive oxygen species (e.g. Clare et al., 2008). XOD exists in two separate but interconvertible forms, xanthine dehydrogenase and xanthine oxidase. In dehydrogenase catalysed reactions formation of NADH is favoured, while oxidase catalysis converts oxygen into reactive oxygen species. The conversion process has been described in quite a few publications (e.g. Rasmussen et al., 2000; Nishino et al., 2008).

Figure 3. Schematic presentation of the location of cysteines in bovine xanthine oxidoreductase (Rasmussen et al., 2000). Positions and residue numbers of labelled cysteines are illustrated, as well as the cysteines involved in binding of the two 2Fe/2S centres. Cys535, 992, 1317, and 1325 are of special interest for conversion (Nishino et al., 2008). Proposed triple domain structure of the enzyme is depicted. Click to enlarge figure.
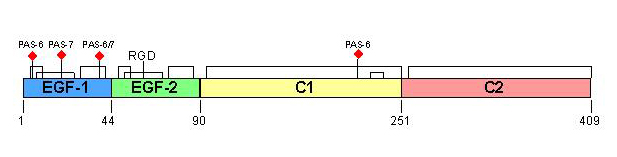
Lactadherin (synonymous with PAS-6/7, bovine-associated mucoprotein, BA-46, P47 or MFG-E8) is a widely distributed glycoprotein (~ 50 kDa) that was originally characterized by its association with milk fat/lipid globule membranes (Hvaaregaard et al., 1996).
Our experiments have shown that lactadherin can act as a link between two surfaces by binding to integrin receptors (?v?3 and ?v?5) through its N-terminus and to phospholipids through its C-terminus (Andersen et al., 1997; Andersen et al., 2000). Additional studies have shown that lactadherin showed preferential binding to phosphatidylserine (PS). This binding was shown to take place in a Ca++-independent manner, to be stereoselective for the L-form, and that lactadherin binding is enhanced by membrane curvature (Shi et al., 2004; Shao et al., 2008). This prompted us in year 2000 to suggest that “Since apoptosis-derived bodies display PS on their surface, lactadherin may be important for the macrophage uptake of these bodies” and that “a model in which PS-associated lactadherin binds to e.g. macrophage integrin receptors prior to endocytosis may be interesting to pursue” (Andersen et al., 2000). Shortly after this function was actually demonstrated in vivo (Hanayama et al., 2004).
During the following decade lactadherin was recognized to be a multifunctional protein participating in various processes, e.g. opsonized clearance of apoptotic cells by macrophage phagocytosis, inflammation, adult onset-autoimmune lupus-like pathology, spenomegaly, and defective germinal center formation. Relations for lactadherin to sickle cell anemia, Alzheimers’s disease, neovascularization and angiogenesis have also been reported. In the mammary gland, lactadherin have been suggested to be important for secretion of milk fat globules and the removal of these during involution. Finally, milk lactadherin has been shown to hold attenuating activities against rotavirus and enterotoxic E. coli. (for references see Rasmussen et al, 2009; Raymond et al, 2009).
We have utilized the inherent ability of lactadherin to recognize PS in studies validating its potential as an in vitro imaging agent of apoptosis. This was done by conjugating purified lactadherin with fluorescent probes or hydrazinonicotinamide (HYNIC) and 99mTc. Activity and selectivity for PS has been estimated on apoptotic suspension cell lines (HL60 and K562) by means of flow cytometry, confocal microscopy for fluorescein, and more conventional binding assays. The performance of lactadherin has been compared with annexin V (the bench-mark PS-probe) in several studies. All performed experiments indicate that the lactadherin conjugates are indeed stable and biologically active with a specific affinity for PS containing membranes (Shi et al., 2004, Shi et al, 2006; Shi et al, 2006; Waehrens et al., 2007; Waehrens et al., 2009; Faldborg et al., 2010 a.o.).
Andersen, M.H., Berglund, L., Rasmussen, J.T. and Petersen, T.E. (1997), Bovine PAS-6/7 binds alpha-v-beta-5 integrin and anionic phospholipids through two domains, Biochemistry 18, 5441-5446.
Andersen, M.H., Graversen, H., Fedosov, S.N., Petersen, T.E. and Rasmussen, J.T. (2000), Functional analyses of two cellular binding domains of bovine lactadherin, Biochemistry 39, 6200-6206.
Berglund, L., Rasmussen, J.T., Andersen, M.D., Rasmussen, M.S. and Petersen, T.E. (1996a), Purification of the bovine xanthine oxidoreductase from milk fat globule membranes and cloning of complementary deoxyribonucleic acid, J. Dairy Sci. 79, 198-204.
Cavaletto. M., Giuffrida, M.G., and Conti A. (2008), Milk fat globule membrane components - A proteomic approach. Adv. Exp. Med. Biol. 606, 129-141.
Clare, D.A., Zheng, Z., Hassan, H.M., Swaisgood, H.E., Catignani, G.L. (2008), Antimicrobial properties of milkfat globule membrane fractions, J. Food Prot. 71, 126-133.
Dewettinck, K., Rombaut, R., Thienpont, N., Le, T.T., Messens, K. and van Camp., J. (2008), Nutritional and technological aspects of milk fat globule membrane material. Int. Dairy J. 18, 436-457.
Falborg, L., Waehrens, L.N., Alsner, J., Bluhme, H., Frøkiær, J., Heegaard, C.W., Horsman, M.R., Rasmussen, J.T., and Rehling, M. (2010), Biodistribution of 99mTc-HYNIC-Lactadherin in Mice – a Potential Tracer for Visualizing Apoptosis In Vivo. Scand. J. Clin. & Lab. Invest. 70, 209-216.
Hanayama, R., Tanaka, M., Miyasaka, K., Aozasa, K., Koike, M., Uchiyama, Y., and Nagata, S. (2004). Autoimmune disease and impaired uptake of apoptotic cells in MFG-E8-deficient mice. Science 304, 1147-1150.
Hvarregaard, J., Andersen, M.H., Berglund, L., Rasmussen, J.T. and Petersen, T.E. (1996), Characterization of glycoprotein PAS-6/7 from membranes of bovine milk fat globules, Eur. J. Biochem. 240, 628-636.
Mather, I. H. (2000). A review and proposed nomenclature for major proteins of the milk-fat globule membrane. J. Dairy Sci., 83, 203–247.
Nishino, T., Okamoto, K., Eger, B.T., Pai, E.F. and Nishino, T. (2008), Mammalian xanthine oxidoreductase - mechanism of transition from xanthine dehydrogenase to xanthine oxidase, FEBS J. 275, 3278-3289.
Pallesen, L.T., Andersen, M.H., Nielsen, R.L., Berglund, L., Petersen, T.E. Rasmussen, L.K. and Rasmussen, J.T. (2001), Purification of MUC1 from bovine milk-fat globules and characterization of a corresponding full-length cDNA clone, J. Dairy Sci. 84, 2591-2598.
Pallesen, L.T., Berglund, L., Rasmussen, L.K., Petersen, T.E. and Rasmussen, J.T. (2002), Isolation and characterization of MUC15, a novel cell membrane-associated mucin, Eur. J. Biochem, 269, 2755-2763.
Pallesen, L.T., Pedersen, L.R.L. Petersen, T.E., and Rasmussen, J.T. (2007), Characterization of Carbohydrate Structures of Bovine MUC15 and Distribution of the Mucin in Bovine Milk, J. Dairy Sci. 90, 3143-3152.
Pallesen, L.T., Pedersen, L.R.L. Petersen, T.E., Knudsen., C.R. and Rasmussen, J.T. (2008). Characterization of Human MUC15 and Identification of Ovine and Caprine Orthologues, J. Dairy Sci. 91, 4477-4483
Parker, P., Sando, L., Pearson, R., Kongsuwan, K., Tellam, R.L., and Smith, S. (2010), Bovine Muc1 inhibits binding of enteric bacteria to Caco-2 cells. Glycoconj. J. 27, 89-97.
Patton, S., S. J. Gendler, and A. P. Spicer. (1995), The epithelial mucin, MUC1, of milk, mammary gland and other tissues, Biochim. Biophys. Acta 124, 407-424.
Patton, S. (1999), Some practical implications of the milk mucins, J. Dairy Sci. 82, 1115-1117.
Puddu, P., Puddu, G.M., Cravero, E., Rosati, M., Muscari, A. (2008), The molecular sources of reactive oxygen species in hypertension, Blood Press. 17, 70-77.
Raymond, A., Ensslin, M.A., and Shur, B.D. (2009), SED1/MFG-E8: a bi-motif protein that orchestrates diverse cellular interactions. J. Cell. Biochem. 106, 957-966.
Rasmussen, J. T. (2009), "Bioactivity of milk fat globule membrane proteins", Aust. J. Dairy Technol. 64, 63-67.
Rasmussen, J. T., Berglund, L., Pallesen, L. T., & Petersen, T. E. (2002), Proteins from the milk fat globule membrane. Poster at the 26th IDF World Dairy Congress, September 24–27, Paris, France.
Rasmussen, J.T., Rasmussen, M.S. and Petersen, T.E. (2000), Cysteines involved in the interconvertion between dehydrogenase and oxidase forms of bovine xanthine oxidoreductase, J. Dairy Sci. 83, 499-506.
Shao, C., Novakovic, V.A., Head, J.F., Seaton, B.A. and Gilbert, G.E. (2008), Crystal structure of lactadherin C2 domain at 1.7A resolution with mutational and computational analyses of its membrane-binding motif, J. Biol. Chem. 283, 7230-7241
Shi, J., Heegaard, C.W., Rasmussen, J.T. and Gilbert, G.E. (2004), Lactadherin binds selectively to membranes containing phosphatidyl-L-serine and curvature strain. Biochim. Biophys. Acta, 1667, 82-90.
Shi, J., Shi, Y., Waehrens, L.N., Rasmussen, J.T., Heegaard, C.W., and Gilbert, G.E. (2006), Lactadherin detects early phosphatidylserine exposure on immortalized leukaemia cells undergoing programmed cell death, Cytometry A. 69, 1193-1201.
Waehrens, L.N., Heegaard, C.W., Gilbert, G.E., and Rasmussen, J.T. (2009) Bovine Lactadherin as a Calcium-independent Imaging Agent of phosphatidylserine Expressed on the Surface of Apoptotic HeLa Cells. J. Hist. Cyt., 10, 907-914.
Waehrens, L.N., Rasmussen, J.T, Heegaard, C.W., and Falborg, L. (2007), Preparation and in vitro evaluation of 99mTc-labelled bovine lactadherin as a novel radioligand for apoptosis detection. J. Label. Compd. Radiopharm. 50, 211–217.
Zhang, J., Perez, A., Yasin, M., Soto, P., Rong, M., Theodoropoulos, G., Carothers Carraway, C.A. and Carraway, K.L. (2005), Presence of MUC4 in human milk and at the luminal surfaces of blood vessels, J. Cell Physiol. 204, 166-177.